100年9月(第231期)
植物對環境逆境之調控與應用
農業藥物毒物試驗所 蔣永正
一 . 前言
作物生育過程的每一階段都受到環境的影響與控制。植物和動物不同,無法自由移動尋找合適的環境,面對會 影響生長或改變生理特性的逆境時 , 為了生存必須發展出適應 環境的能力。環境中如日照強度、溫度、雨量、鹽分濃度、重金屬、除草劑及空氣、水等污染引起之 異常 ,往往 會促使植體內自由基及活性氧的產生。一般 氧氣在參與細胞新陳代謝的過程中,容易被活化成為具強氧化力的活性氧。正常情況下,植物體內活性氧的產生和清除是處於動態平衡狀態。但逆境會導致抗氧化系統對活性氧的清除能力下降,發生氧化傷害,造成代謝功能的不可修復和細胞死亡。本文即針對植物的抗氧化系統,對環境逆境產生之活性氧化物的反應差異,藉以探討植物對逆境的調適作用及應用之可能性。
二 . 植物體活性氧化物( reactive oxygen species, ROS )的發生與作用
氧氣為 植物生命活動中不可缺少的重要分子 。一般在基底態之氧分子活性低,對有機物不會造成危害,但是活性氧化物則對細胞有毒害作用。當葉綠體光反應中心截取光能後,正常狀況下會產生氧氣及還原態的 NADPH ,提供 CO2 固定所需之基質,但當 PS I 還原端之電子被帶到氧分子上,取代原來正常還原 NADP+ 的路徑,則會產生活性氧
|
植物細胞內許多正常進行的代謝反應;如光合作用及呼吸作用,均會產生不同種類的活性氧化物(表 1 );非生物性逆境引起之代謝路徑中,也會如位於過氧化體內之 glycolate oxidase ,在光呼吸過程中產生 H2O2 。此外 NADPH oxidase 、 amine oxidase 及 cell wall-bound peroxidases ,亦為促使活性氧化物產生的途徑。
植物在正常生長狀況下,細胞中活性氧化物的產生量很低( |
活性氧化物導致細胞死亡的途徑為膜脂的過氧化、蛋白質氧化、酵素活性抑制及 DNA 與 RNA 受損。但逆境期間促使活性氧化物產生,雖然對細胞造成威脅,卻也具有啟動逆境防禦系統的訊息傳導( signal transduction )功能;如活化細胞的死亡( programmed cell death, PCD )機制,即由過氧化逆境誘導細胞局部死亡。活性氧化物若被視作細胞逆境的監測指標時,其在細胞內的含量則需嚴格控制,因為在本身有毒但又具有參與訊息傳導功能的雙重角色下,需有適合之機制來調控細胞內的濃度。除了能夠微調活性氧化物至低量以達到訊息傳導的目的,又能夠將超量之活性氧化物解毒,以延續生物在逆境中之存活。
三 . 植物對氧化逆境之防禦系統
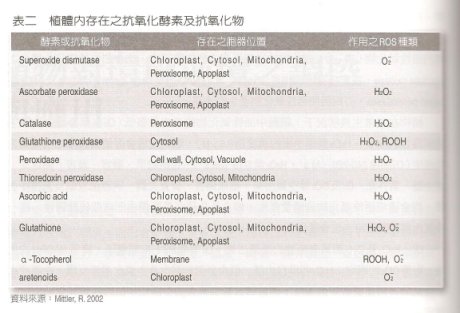
植物體清除活性氧的保護系統又稱為抗氧化系統, 包括抗氧化酵素及非酵素型抗氧化 劑 (表 2 ),以應付具擴散性、水溶性及可與各種生物分子反應之活性氧化物,避免發生使活性物種毒性加強的連鎖反應。
(一)抗氧化酵素 清除系統
在 酵素抗氧化系統中 包括對活性氧直接作用的超氧歧化酶( SOD )、過氧化氫酶( catalase, CAT )、過氧化酶( peroxidase, POD ),以及保持抗氧化物質還原性的 ascorbate-glutathione cycle 中的抗壞血酸過氧化酶( ascorbate peroxidase, APX )、穀胱苷肽還原酶( glutathione reductase, GR )、及單脫氫抗壞血酸還原酶( monodehydroascorbate reductase, MDHAR )等。
1. 超氧歧化酶( s uperoxide dismutase, SOD )
主要的作用是將 ![]() 轉換成 H2O2 ,為防禦系統的第一線,進行之歧化作用會被金屬離子所催化。具有 Mn-SOD 、 Fe-SOD 及 Cu/Zn-SOD 三種 同功酶 ,分別位於粒線體、葉綠體及 細胞質 ,且均由核基因所控制。在 菸草葉片中的 SOD 同功酶形式受發育階段的調控,幼葉中以 Cu/Zn-SOD 較多,活性隨葉齡增加而下降;老葉中主要為 Fe-SOD 和 Mn-SOD 。 SOD 的活性由
轉換成 H2O2 ,為防禦系統的第一線,進行之歧化作用會被金屬離子所催化。具有 Mn-SOD 、 Fe-SOD 及 Cu/Zn-SOD 三種 同功酶 ,分別位於粒線體、葉綠體及 細胞質 ,且均由核基因所控制。在 菸草葉片中的 SOD 同功酶形式受發育階段的調控,幼葉中以 Cu/Zn-SOD 較多,活性隨葉齡增加而下降;老葉中主要為 Fe-SOD 和 Mn-SOD 。 SOD 的活性由 ![]() 和 H2O2 的濃度決定。逆境下細胞中的活性氧分子會升高, SOD 活性的表現也會增加。 SOD 在不同逆境下的調控及反應有差別,顯示與植體對逆境的忍受性有關。
和 H2O2 的濃度決定。逆境下細胞中的活性氧分子會升高, SOD 活性的表現也會增加。 SOD 在不同逆境下的調控及反應有差別,顯示與植體對逆境的忍受性有關。
2. 過氧化氫酶( catalase, CAT )與過氧化酶( peroxidase, POD )
CAT 對 H2O2 具專一性, 可直接將其轉變為 H2O ,對氧化逆境有極佳之保護作用。 主要存在於過氧化體中,負責清除過氧化體中產生的 H2O2 。乙醛酸體中也發現有 CAT 的存在,主要是清除光呼吸過程或脂肪酸 β- 氧化反應形成的 H2O2 。由於 H2O2 可直接跨膜擴散,其他部分產生的 H2O2 也可擴散到過氧化體中被 CAT 分解,與 SOD 協力清除植體內潛在危害的![]() 和 H2O2 ,將 OH- 發生的機率降至最低。
和 H2O2 ,將 OH- 發生的機率降至最低。
POD 廣泛存在於植物體內的不同組織,屬活性較高的誘導酶,能夠反應植物的代謝狀況和對環境的適應性。 POD 具有雙重的作用特性,在逆境或衰老初期的表現為清除 H2O2 ,達到保護細胞的功能。但在逆境或衰老後期,則參與活性氧的生成、葉綠素的降解,引發膜脂過氧化作用,加速細胞的衰老死亡,成為植物的衰老指標。
3. 抗壞血酸過氧化酶( ascorbate peroxidase, APX )、穀胱苷肽還原酶( glutathione reductase, GR )、及單脫氫抗壞血酸還原酶( monodehydroascorbate reductase, MDHAR ) APX 、 GR 及 MDHAR 為葉綠體內 ascorbate-glutathione cycle 內的主要酵素。 APX 為此循環的第一個酵素,可將 H2O2 還原成水,同時氧化 ascorbate 成為單脫氫抗壞血酸( monodehydroxyascorbate, MDA ), MDA 與 ferredoxins 的非酵素反應,或利用 NADPH 在 MDAR 催化下可再還原成 ascorbate 。 APX 在葉綠體內具有與葉綠體基質或類囊膜結合的兩種型式,另外還有細胞質同功酶。葉綠體內清除活性氧化物的作用主要發生在類囊膜表面,接近 PS I ,以防止活性氧化物逃脫或互相反應,為防禦的第二道防線,以保護 Calvin cycle 中的敏感酵素受到攻擊。
GR 則以 NADPH 作為電子授與者,將 GSSG 還原成 GSH ,完成了 ascorbate-glutathione cycle 的反應。 GR 活性 80% 都在葉綠體內,粒線體及 cytosol 中也有部分同功酶。 GR 及 APX 對逆境反應的活性變化頗分歧,與植物種類及活性氧化物來源都有關係。
細胞內 SOD 及 APX 或 CAT 間的平衡,可決定 ![]() 及 H
2O
2 的濃度,此種平衡也包括金屬離子的隔絕,防止 Fenton reaction 的發生及生成 OH
- 。此外由 APX 及 CAT 與 H
2O
2 的親和力高低可知; APX ( μM )應負責活性氧化物訊息傳遞之微細調節, CAT ( mM )則負責逆境下移除過量產生之活性氧化物。
及 H
2O
2 的濃度,此種平衡也包括金屬離子的隔絕,防止 Fenton reaction 的發生及生成 OH
- 。此外由 APX 及 CAT 與 H
2O
2 的親和力高低可知; APX ( μM )應負責活性氧化物訊息傳遞之微細調節, CAT ( mM )則負責逆境下移除過量產生之活性氧化物。
(二)抗氧化劑 清除系統
主要指抗壞血酸( ascorbate, ASA )、類胡蘿蔔素( carotenoids )及一些含硫基的低分子化合物,如還原型 穀胱苷肽( reduced glutathione, GSH )等,它們通過不同途徑直接或間接地將活性氧清除。
1. 抗壞血酸( ascorbate, ASA )
ASA 可作為 APX 的反應基質,清除葉綠體內的 H 2O 2 ,同時在 類囊膜表面可保護或再生氧化態之 carotenes 及 tocopherols 。但濃度過高會將 Fe +3 還原成 Fe +2 , 反而造成 OH - 的產生。
ASA 存在於葉綠體基質中,是 ![]() 和 OH
- 的有效清除劑,同時也是
和 OH
- 的有效清除劑,同時也是 ![]() 的 淬滅 劑( quencher )。 ASA 可以清除膜脂過氧化過程中產生的 多聚不飽和脂肪酸( PUFA )自由基。它不僅作為 APX 的反應基質,還可以當 作抗氧化劑直接清除活性氧。 ASA 可還原
的 淬滅 劑( quencher )。 ASA 可以清除膜脂過氧化過程中產生的 多聚不飽和脂肪酸( PUFA )自由基。它不僅作為 APX 的反應基質,還可以當 作抗氧化劑直接清除活性氧。 ASA 可還原 ![]() ,清除 OH
- , 淬滅
,清除 OH
- , 淬滅 ![]() ,歧化 H
2O
2 ,還可再生 tocopherol 。由於 ASA 具有多種抗氧化功能,當 ASA 含量降低時,顯示植物抗氧化能力的整體衰退。
,歧化 H
2O
2 ,還可再生 tocopherol 。由於 ASA 具有多種抗氧化功能,當 ASA 含量降低時,顯示植物抗氧化能力的整體衰退。
2. 類胡蘿蔔素( carotenoids )
類胡蘿蔔素 會消除葉綠素的激化狀態及照光後產生之![]() ,免除脂質的過氧化作用;激化之 carotene 則將能量傳遞至其他的天線色素,或以熱的型式消散而回到穩定的基底態。
,免除脂質的過氧化作用;激化之 carotene 則將能量傳遞至其他的天線色素,或以熱的型式消散而回到穩定的基底態。
類胡蘿蔔素存在於葉綠體內,一方面阻止激發態葉綠素分子的激發能從反應中心向外傳遞;另一方面,它也保護葉綠素分子免於遭受光氧化傷害。類胡蘿蔔素共有 α-carotene 、 β-carotene 及葉黃素三種形式,以 β-carotene 含量最高,為 ![]() 最有效的 淬 滅劑。但 β-carotene 的抗氧化作用受氧濃度的影響,低氧下有良好的抗氧化作用,高氧下則會轉化成自由基的形式,加速氧化的進程。
最有效的 淬 滅劑。但 β-carotene 的抗氧化作用受氧濃度的影響,低氧下有良好的抗氧化作用,高氧下則會轉化成自由基的形式,加速氧化的進程。
3. 還原型 穀胱苷肽( reduced glutathione, GSH )
GSH 可有效還原或解毒許多氧化物種,本身可利用 NAD ( P ) H 再生,對與抗氧化系統有關之 ascorbate 或 tocopherol 的再循環利用十分重要,通常在逆境下 glutathione pools 有減少的趨勢。 GSH 不僅參與 ascorbate 的再生( ascorbate-glutathione cycle ),同時還可與![]() 和 OH
- 等活性氧化物作用以保護蛋白質。
和 OH
- 等活性氧化物作用以保護蛋白質。
一般認為氧化型和還原型穀胱苷肽的總量下降,但 GSH/GSSG 比值高時,抗氧化保護作用較佳。但也有研究指出, GSH 大量增加,反而可能造成更大程度的氧化逆境。
GSH 可與包括 OH - 在內的多種自由基直接發生反應,並可將有機自由基 R 還原為 RH 。還原型的穀胱苷肽可將系統中產生的雙脫氫抗壞血酸( DHA )還原成 ASA , GSSG 在 GR 作用下又還原成 GSH , GSH 還能與 α-tocopherol 協同作用,清除細胞內累積的自由基。
Ascorbate 及 glutathione 等抗氧化物在葉綠體內的濃度很高,兩者在細胞內的含量比為決定活性氧化物是否需要清除的依據。不同抗氧化物的氧化態與還原態比值(如 GSSG/GSH ),也能夠作為調節活性氧化物清除機制之訊息。
4. 生育酚( t ocopherol )
最主要的 isomer 為 α-tocopherol ,在終結自由基之連鎖反應上扮演重要角色。葉綠體膜系的脂質不飽和作用也需要大量的 α-tocopherol 參與。 α-tocopherol 是膜脂過氧化的有效抑制劑,其含量的增加,可能表示植物膜系對大量產生自由基的保護反應。
此外,苯甲酸、異丙醇可直接清除 OH
- ,多酚、單甯與黃酮類等二次代謝物可直接清除 ![]() ,沒食子酸丙酯清除
,沒食子酸丙酯清除 ![]() 的能力與 SOD 十分接近。鋅含量高時,芥菜和綠豆體內自由基的產生增加,同時脯氨酸( proline )也大量累積。在分離的粒線體中,證實脯氨酸可減少高光照所誘導的自由基發生量。推測脯氨酸的累積與自由基的清除有關。其他如山梨醇( sorbitol )與甘露醇( mannitol )等滲透調節物質也具有清除活性氧,抑制膜脂過氧化的能力。
的能力與 SOD 十分接近。鋅含量高時,芥菜和綠豆體內自由基的產生增加,同時脯氨酸( proline )也大量累積。在分離的粒線體中,證實脯氨酸可減少高光照所誘導的自由基發生量。推測脯氨酸的累積與自由基的清除有關。其他如山梨醇( sorbitol )與甘露醇( mannitol )等滲透調節物質也具有清除活性氧,抑制膜脂過氧化的能力。
針對細胞能夠感應不同胞器發生之氧化逆境,及逆境訊息傳遞過程的研究,發現一種在各類逆境下釋出之荷爾蒙( phytohormones ),可能擔負此種生理上的調節作用,即荷爾蒙在細胞面臨氧化還原的變化時,率先啟動防禦系統,將訊息傳達至不同的組織,且維持一段可供防禦機制啟動的較長期間。
四 . 植物 抗氧化系統對逆境之反應
抗氧化酵素及抗氧化劑分子在細胞內都有一定的含量水準,以應付正常代謝作用產生之活性物,或是突發逆境所誘導的毒害。以下就乾旱、重金屬及除草劑為例,說明 逆境下之 植物 抗氧化酵素或抗氧化分子的變化反應。
(一)乾旱
棉花及 甘薯 等作物在輕微缺水下, SOD 活性上升,但 超過一定程度, 活性則下降。不同品種之 SOD 活性對 乾旱 的敏感程度有差別。耐旱性越強之雲薹品種,隨乾旱時間延長, SOD 活性降低較慢。玉米不同生育期遭受到 乾旱時 , SOD 活性均降低。
水稻不同品種之 POD 及 CAT 活性對乾旱的反應不同。 CAT 活性均在輕度缺水時升高,重度乾旱時又下降,不同品種間之變化幅度不同。甘薯不論品種耐旱與否,隨缺水程度加深, POD 活性下降, CAT 活性呈升高趨勢。冬小麥在土壤缺水初期,劍葉的 AP 和 GR 活性均呈現先上升、後下降的變化趨勢,抗旱性強的品種 AP 活性變化與此相同,但 GR 活性卻持續上升。 顯示在乾旱缺水下,一些植物葉綠體中的 H2O2 是經由抗氧化酵素反應清除的,另一些植物中的 H2O2 則可能是由其他途徑予以清除。
綜合上述試驗 結果,植物抗氧化酵素對乾旱逆境的反應,可能由其本身所感受到的水分缺少程度所啟動。通常發生輕中度之缺水逆境時,抗氧化酵素的活性增加,活性氧造成的傷害得以緩解。
小麥在輕度缺水時,耐旱品種的 ASA 和 GSH 含量變化不大。但在嚴重乾旱下,兩者含量會迅速下降,恢復供水後耐旱品種雖有所回升,但與對照處理有明顯差異。 ASA 含量的下降,會直接導致活性氧清除能力的降低。以 30%PEG 模擬嚴重乾旱之環境條件,造成水稻膜脂過氧化及葉片白化現象,同時葉片中 ASA 、 GSH 、 α-tocopherol 及 β- carotene 等抗氧化劑均顯著減少。但乾旱對豌豆的 ASA 含量無明顯影響,卻引起 β- carotene 含量增加 18% , α-tocopherol 增加 66% 。
(二)重金屬
|
O2 α-tocopherol 是膜脂過氧化的有效抑制劑,其含量的增加,可能表示植物膜系對大量產生的自由基的保護反應。 |
|
O2 α-tocopherol 是膜脂過氧化的有效抑制劑,其含量的增加,可能表示植物膜系對大量產生的自由基的保護反應。 |
銅離子會誘導脂質氧化酶( lipoxygenase )的活化,啟動脂質過氧化,減少葉綠素和血紅素 heme 含量,造成葉片白化現象。 營養液中 Cd 含量在 1 ppm 時,能顯著增加白菜 POD 、 CAT 及 APX 活性,當增加至 10 ppm 時卻會明顯降低 SOD 、 POD 、 CAT 及 APX 活性,促使脫氫抗壞血酸還原酶和穀胱苷肽還原酶活性增加。
鉛對小麥 POD 具有啟動效應,且隨逆境程度的增加,效應越明顯。對 SOD 和 CAT 在逆境初期的表現為啟動效應,後期隨程度加重則為抑制效應,顯示 POD 在鉛引起之抗逆境反應中扮演關鍵角色。
(三) 除草劑
聯吡啶類( bipyridinium )除草劑巴拉刈 光照下在葉綠體內,經由連鎖反應會不斷的產生 ![]() ,再由 SOD 的作用生成 H
2O
2 , H
2O
2 則由 APX 及 GR 在 ascorbate-glutathione cycle 內催化 ascorbate 及 glutathione 的氧化及再還原反應,變成 H
2O 移出。在水稻切離葉片內發現,巴拉刈引起脂質過氧化毒害時,會伴隨 APX 、 GR 及 CAT 活性的降低, SOD 活性則在毒害發生至某一程度才會降低,但細胞內 H
2O
2 含量則無明顯變化,推測可能用為參與 Fenton reaction 產生 OH
- ;此外 GSH 等自由基清除劑以外施方式,可明顯抑制巴拉刈引起之脂質過氧化作用,及增加 CAT 及 POD 等抗氧化酵素活性。因此抗氧化酵素活性及擔任自由基清除劑之抗氧化物,與巴拉刈導致之脂質過氧化毒害程度有關。
,再由 SOD 的作用生成 H
2O
2 , H
2O
2 則由 APX 及 GR 在 ascorbate-glutathione cycle 內催化 ascorbate 及 glutathione 的氧化及再還原反應,變成 H
2O 移出。在水稻切離葉片內發現,巴拉刈引起脂質過氧化毒害時,會伴隨 APX 、 GR 及 CAT 活性的降低, SOD 活性則在毒害發生至某一程度才會降低,但細胞內 H
2O
2 含量則無明顯變化,推測可能用為參與 Fenton reaction 產生 OH
- ;此外 GSH 等自由基清除劑以外施方式,可明顯抑制巴拉刈引起之脂質過氧化作用,及增加 CAT 及 POD 等抗氧化酵素活性。因此抗氧化酵素活性及擔任自由基清除劑之抗氧化物,與巴拉刈導致之脂質過氧化毒害程度有關。
巴拉刈處理抗、感野茼蒿(Conyza sumatrensis )後,以葉綠素螢光 Fv/Fm 達 50% 抑制計算其抗感比值,當抗感比大時, SOD 及 GR 活性均增高,抗感比相差小時,兩者的酵素活性也相當。巴拉刈處理之抗、感加拿大蓬(C .canadensis )植株,於噴施後 2 小時,抗性株內 APX 活性下降, 24 小時後開始回升; GR 及 SOD 活性分別增加約 20-25% 及 58% ;配合葉綠素螢光 Fv/Fm 計算之抗感比卻在 450-1000 範圍;顯示抗氧化酵素活性的增加程度與抗感比間似無明顯相關,即此種與生俱來之保護機制似不足以消除連續產生的過氧化物。從上述不同試驗結果顯示;細胞抗氧化酵素活性及抗氧化物含量,與巴拉刈在植體造成之氧化毒害程度有關,但是否為構成抗性的主要機制,尚須進行系統研究以提供足以研判之完整數據。
五 . 結語
植物對逆境的抗性與特定酵素或同功酶的表現與啟動有關。由植物抗氧化 酵素活性或抗氧化劑含量 對逆境壓力反應的 變化 , 可應用為栽培環境中異常因子發生的偵測指標。 但因為逆境類別、強度與曝露期長短等的不同,及作物品種、生育期與酵素類型的差異,導致植物對逆境壓力的生理反應極為複雜,造成試驗結果的分歧及缺乏可供應用的一致性結論。有關植物抗氧化系統對逆境壓力的反應,雖已累積可觀的研究資料,但針對抗氧化酵素在逆境壓力中活性變化的機制、作物品種間反應差異的原因、抗氧化劑及滲透調節物質的作用途徑與關鍵步驟等尚須深入探討,才可進一步利用為提高作物抗性能力的準確依據。
表 1 植體內發生的活性氧化物( Reactive oxygen species, ROS )
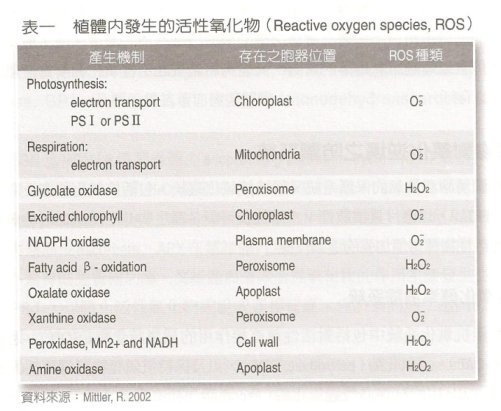
表 2 植體內存在之抗氧化酵素及抗氧化物